

Benefits of whole ginger extract in prostate cancer
- PMID: 21849094
- PMCID: PMC3426621
- DOI: 10.1017/S0007114511003308
Benefits of whole ginger extract in prostate cancer
Abstract
It is appreciated far and wide that increased and regular consumption of fruits and vegetables is linked with noteworthy anticancer benefits. Extensively consumed as a spice in foods and beverages worldwide, ginger (Zingiber officinale Roscoe) is an excellent source of several bioactive phenolics, including non-volatile pungent compounds such as gingerols, paradols, shogaols and gingerones. Ginger has been known to display anti-inflammatory, antioxidant and antiproliferative activities, indicating its promising role as a chemopreventive agent. Here, we show that whole ginger extract (GE) exerts significant growth-inhibitory and death-inductory effects in a spectrum of prostate cancer cells. Comprehensive studies have confirmed that GE perturbed cell-cycle progression, impaired reproductive capacity, modulated cell-cycle and apoptosis regulatory molecules and induced a caspase-driven, mitochondrially mediated apoptosis in human prostate cancer cells. Remarkably, daily oral feeding of 100 mg/kg body weight of GE inhibited growth and progression of PC-3 xenografts by approximately 56 % in nude mice, as shown by measurements of tumour volume. Tumour tissue from GE-treated mice showed reduced proliferation index and widespread apoptosis compared with controls, as determined by immunoblotting and immunohistochemical methods. Most importantly, GE did not exert any detectable toxicity in normal, rapidly dividing tissues such as gut and bone marrow. To the best of our knowledge, this is the first report to demonstrate the in vitro and in vivo anticancer activity of whole GE for the management of prostate cancer.
Conflict of interest statement
The authors declare that they have no conflict of interest.
Figures
 ), DU145 (
), DU145 (
 ), PC-3 (
), PC-3 (
 ), C4-2 (
), C4-2 (
 ), C4-2B (
), C4-2B (
 ) cells, as well as normal prostate epithelial cells (PrEC,
) cells, as well as normal prostate epithelial cells (PrEC,
 ) and human dermal primary fibroblasts (HDF,
) and human dermal primary fibroblasts (HDF,
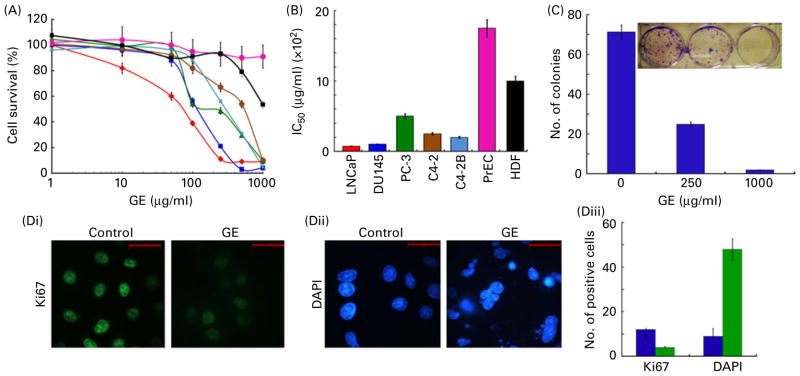
 ) were treated with gradient concentrations of GE for 72 h. The percentage of cell proliferation at indicated concentrations, compared with untreated control cells, was measured by the in vitro cell proliferation assay, as described in Materials and methods. (A) Plot of percentage of cell survival vs. GE concentrations used for the determination of half-maximal concentration of growth inhibition (IC50) values. Values are means of three independent experiments performed in triplicate, with standard deviations represented by vertical bars (P<0·05). (B) Bar graph representation of the IC50 of the indicated cell lines. (C) Bar graph representation and photograph of crystal violet-stained surviving colonies from the control and GE-treated (250 and 1000 μg/ml) groups. (D) Fluorescence micrographs of control and GE-treated PC-3 cells stained for (Di) Ki67 (green) or (Dii) 4′,6-diamidino-2-phenylindole (DAPI, blue). Scale bar, 20 μm. (Diii) Quantification of Ki67-postive or DAPI-stained cells in control (
) were treated with gradient concentrations of GE for 72 h. The percentage of cell proliferation at indicated concentrations, compared with untreated control cells, was measured by the in vitro cell proliferation assay, as described in Materials and methods. (A) Plot of percentage of cell survival vs. GE concentrations used for the determination of half-maximal concentration of growth inhibition (IC50) values. Values are means of three independent experiments performed in triplicate, with standard deviations represented by vertical bars (P<0·05). (B) Bar graph representation of the IC50 of the indicated cell lines. (C) Bar graph representation and photograph of crystal violet-stained surviving colonies from the control and GE-treated (250 and 1000 μg/ml) groups. (D) Fluorescence micrographs of control and GE-treated PC-3 cells stained for (Di) Ki67 (green) or (Dii) 4′,6-diamidino-2-phenylindole (DAPI, blue). Scale bar, 20 μm. (Diii) Quantification of Ki67-postive or DAPI-stained cells in control (
 ) and 250 μg/ml of GE-treated PC-3 cells (
) and 250 μg/ml of GE-treated PC-3 cells (
 ) from random image fields totalling 200 cells. Values are means, with standard deviations represented by vertical bars. Mean values were significantly different from the controls (P<0·05). (A colour version of this figure can be found online at
) from random image fields totalling 200 cells. Values are means, with standard deviations represented by vertical bars. Mean values were significantly different from the controls (P<0·05). (A colour version of this figure can be found online at 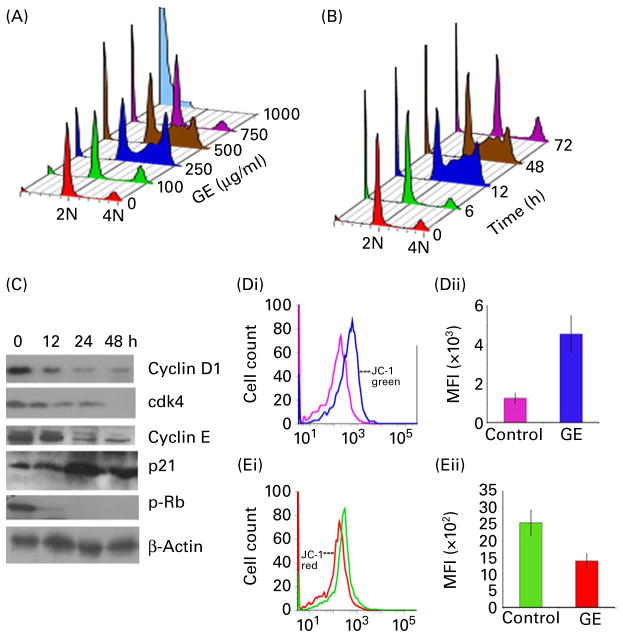
 ) or GE (blue profile;
) or GE (blue profile;
 ) for 24 h. Representative data from a single experiment are shown. (Dii) Quantification of the increase in mean fluorescence intensity (MFI, i.e. the percentage of green JC-1-stained cells) in PC-3 cultures treated with DMSO (control) or GE for 24 h. (Ei) Histogram profiles showing a spectral shift and loss of red fluorescence, consistent with the loss of transmembrane potential on GE treatment (control, green; GE, red). (Eii) Quantification of the decrease in mean fluorescence intensity (i.e. the percentage of red JC-1-stained cells) in PC-3 cultures treated with DMSO (control) or GE for 24 h. Values are means of three independent experiments performed in triplicate, with standard deviations represented by vertical bars (P < 0·05). (A colour version of this figure can be found online at
) for 24 h. Representative data from a single experiment are shown. (Dii) Quantification of the increase in mean fluorescence intensity (MFI, i.e. the percentage of green JC-1-stained cells) in PC-3 cultures treated with DMSO (control) or GE for 24 h. (Ei) Histogram profiles showing a spectral shift and loss of red fluorescence, consistent with the loss of transmembrane potential on GE treatment (control, green; GE, red). (Eii) Quantification of the decrease in mean fluorescence intensity (i.e. the percentage of red JC-1-stained cells) in PC-3 cultures treated with DMSO (control) or GE for 24 h. Values are means of three independent experiments performed in triplicate, with standard deviations represented by vertical bars (P < 0·05). (A colour version of this figure can be found online at 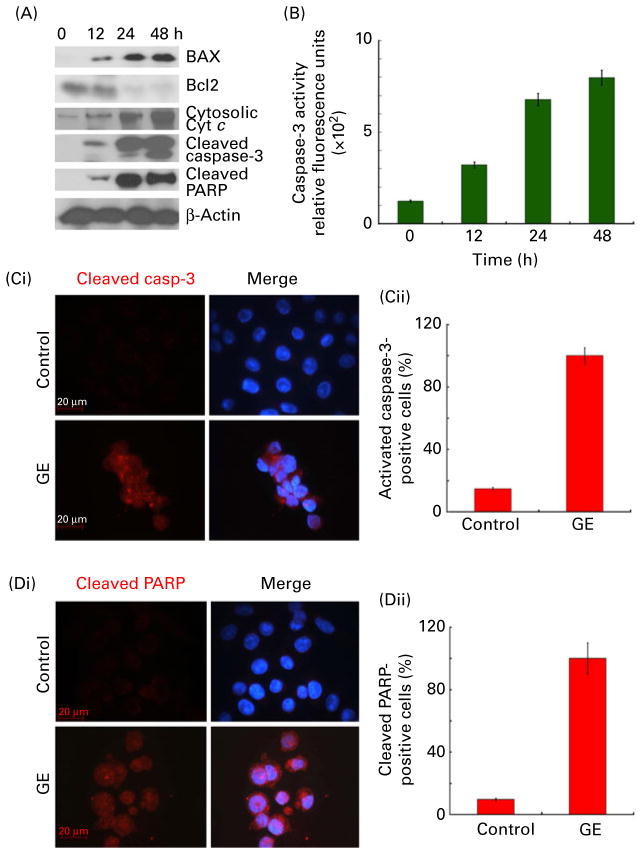
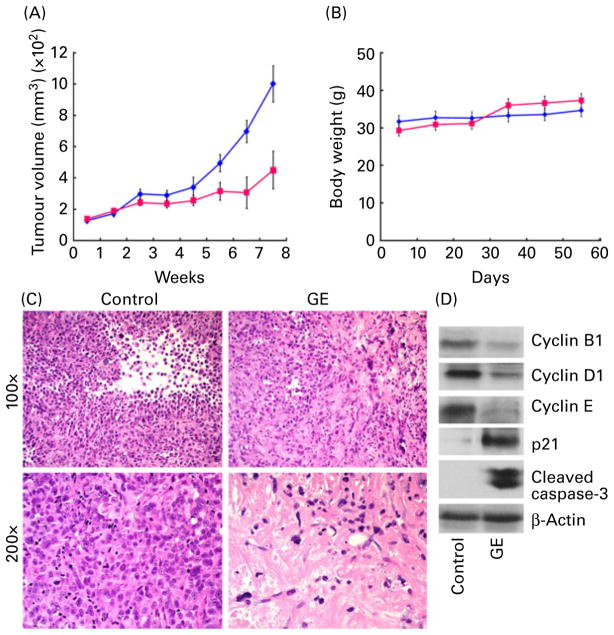
 ) and GE-treated (
) and GE-treated (
 ) mice at the time of treatment. (B) GE treatment was well tolerated, and the body weights of the control (
) and GE-treated (
) groups were comparable. Values are means, with standard deviations represented by vertical bars (n = 6, P < 0·05). (C) Tumour micrographs from control and GE-treated mice, respectively, at 100× and 200× magnification. GE-treated tumour microsections reveal large areas of tumour cell death, consistent with the therapeutic effects of GE. Microsections from control tumour tissue show sheets of tumour cells with high-grade pleomorphic nuclei with minimal cell death. (D) Western blot analysis of tumour tissue lysates from control and GE-treated mice for cyclin B, cyclin D1, cyclin E, p21 and cleaved caspase-3. (A colour version of this figure can be found online at
) mice at the time of treatment. (B) GE treatment was well tolerated, and the body weights of the control (
) and GE-treated (
) groups were comparable. Values are means, with standard deviations represented by vertical bars (n = 6, P < 0·05). (C) Tumour micrographs from control and GE-treated mice, respectively, at 100× and 200× magnification. GE-treated tumour microsections reveal large areas of tumour cell death, consistent with the therapeutic effects of GE. Microsections from control tumour tissue show sheets of tumour cells with high-grade pleomorphic nuclei with minimal cell death. (D) Western blot analysis of tumour tissue lysates from control and GE-treated mice for cyclin B, cyclin D1, cyclin E, p21 and cleaved caspase-3. (A colour version of this figure can be found online at 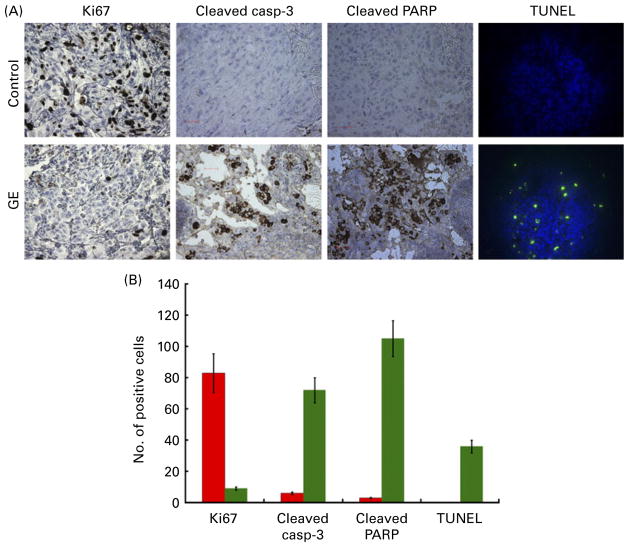
 ; GE,
; GE,
 . (A colour version of this figure can be found online at
. (A colour version of this figure can be found online at Similar articles
-
Enterohepatic recirculation of bioactive ginger phytochemicals is associated with enhanced tumor growth-inhibitory activity of ginger extract.Carcinogenesis. 2014 Jun;35(6):1320-9. doi: 10.1093/carcin/bgu011. Epub 2014 Jan 15. Carcinogenesis. 2014. PMID: 24431413 Free PMC article.
-
Update on the chemopreventive effects of ginger and its phytochemicals.Crit Rev Food Sci Nutr. 2011 Jul;51(6):499-523. doi: 10.1080/10408391003698669. Crit Rev Food Sci Nutr. 2011. PMID: 21929329 Review.
-
Ginger extract adjuvant to doxorubicin in mammary carcinoma: study of some molecular mechanisms.Eur J Nutr. 2018 Apr;57(3):981-989. doi: 10.1007/s00394-017-1382-6. Epub 2017 Feb 22. Eur J Nutr. 2018. PMID: 28229277
-
Ginger phytochemicals exhibit synergy to inhibit prostate cancer cell proliferation.Nutr Cancer. 2013;65(2):263-72. doi: 10.1080/01635581.2013.749925. Nutr Cancer. 2013. PMID: 23441614 Free PMC article.
-
Cross-talk between 10-gingerol and its anti-cancerous potential: a recent update.Food Funct. 2017 Aug 1;8(8):2635-2649. doi: 10.1039/c7fo00844a. Epub 2017 Jul 26. Food Funct. 2017. PMID: 28745358 Review.
Cited by
-
Effect of ginger, chamomile, and green tea extracts on prostate cancer cells.J Genet Eng Biotechnol. 2024 Sep;22(3):100395. doi: 10.1016/j.jgeb.2024.100395. Epub 2024 Jun 19. J Genet Eng Biotechnol. 2024. PMID: 39179318 Free PMC article.
-
Cellular Adaptation Takes Advantage of Atavistic Regression Programs during Carcinogenesis.Cancers (Basel). 2023 Aug 3;15(15):3942. doi: 10.3390/cancers15153942. Cancers (Basel). 2023. PMID: 37568758 Free PMC article. Review.
-
6-shogaol is a potential treatment for Head and Neck Squamous Cell Carcinoma.Int J Med Sci. 2023 Jan 22;20(2):238-246. doi: 10.7150/ijms.80542. eCollection 2023. Int J Med Sci. 2023. PMID: 36794162 Free PMC article.
-
Enterohepatic recirculation of bioactive ginger phytochemicals is associated with enhanced tumor growth-inhibitory activity of ginger extract.Carcinogenesis. 2014 Jun;35(6):1320-9. doi: 10.1093/carcin/bgu011. Epub 2014 Jan 15. Carcinogenesis. 2014. PMID: 24431413 Free PMC article.
-
Molecular Mechanisms and Bioavailability of Polyphenols in Prostate Cancer.Int J Mol Sci. 2019 Mar 1;20(5):1062. doi: 10.3390/ijms20051062. Int J Mol Sci. 2019. PMID: 30823649 Free PMC article. Review.
References
-
- Syed DN, Khan N, Afaq F, et al. Chemoprevention of prostate cancer through dietary agents: progress and promise. Cancer Epidemiol Biomarkers Prev. 2007;16:2193–2203. - PubMed
-
- Nelson WG, De Marzo AM, Isaacs WB. Prostate cancer. N Engl J Med. 2003;349:366–381. - PubMed
-
- Sporn MB. Approaches to prevention of epithelial cancer during the preneoplastic period. Cancer Res. 1976;36:2699–2702. - PubMed
-
- Mann JR, Backlund MG, DuBois RN. Mechanisms of disease: inflammatory mediators and cancer prevention. Nat Clin Pract Oncol. 2005;2:202–210. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
